Garden with Insight v1.0 Help: Plant Biomass Partitioning Model
Note: This was originally written as a design document, so there may be some small
discrepancies between the processes described here and the actual model in Garden with Insight. The
main gist of the submodel is in agreement, though.
First Photosynthate Partitioning - Root, Shoot, Flowers/Fruit, Storage Organs
After floral initiation is complete, flowers will form on the plant (see the section on the reproductive submodel). As the plant creates photosynthate through
photosynthesis, that photosynthate is distributed to the different parts of the plant. There are two stages of
allocation: first to shoot, root and fruit, then to individual shoot meristems and individual fruits. The first
partition is as follows:

Biomass partitioning among roots, shoots, flowers/fruits, and storage organ
The root/shoot ratio is governed by the line between proportion of photosynthate to roots at
germination and that at maturity (EPIC parameters). The only change we are making is that reproductive
and/or storage organ sinks appear as the plant gets older. The strength of the reproductive sink depends
partly on time (as modeled by the increasing proportion) and partly on water stress; as the plant is more
stressed, less photosynthate is allocated to the reproductive biomass. This simulates flower drop and
deficient fruit set in conditions of water stress. So the proportion demanded by the flowers and fruit is
multiplied by the water stress factor to reduce it. To distribute the photosynthate among the four sinks,
they are normalized so they add up to 1.0, then photosynthate is allocated to them in those proportions.
Storage organs can be dealt with in the model in the same way as roots. Storage organs can be in the roots
(sweet potato), in tubers (potato), in the stem (kohlrabi), in axillary buds (Brussels sprouts), in leaf
petioles (celery, onion), and in inflorescences (broccoli, cauliflower). Photosynthate can be allocated to a
storage organ without attention to what plant part(s) the storage organ represents. Therefore allocation to
a storage organ can work in exactly the same way as allocation to fruits -- storage organs demand a
proportion of photosynthate and compete with other plant sinks.
The growth of storage organs occurs almost completely at the end of the life history of most plants and has
a roughly exponential form. We may approximate this by a linear relationship from some starting point to
plant maturity. The starting point of allocation to storage organs is affected by photoperiod in many crops,
but the research on this is not plentiful and there are many exceptions. In this case it is best to base the
starting point of storage allocation simply on HUI. The allocation scheme will look very much like that of
a plant producing fruits (most plants don't produce both at once), with more and more allocation to the
storage organs. Therefore we can simulate a storage organ in the same way as the fruit sink and with
similar parameters. The parameters will be the HUI at the start of storage organ allocation, and the
fraction of photosynthate going to the storage organ at plant maturity (HUI = 1.0).
Storage organs will not be drawn in the sense of growing on the plant. For stems, buds and other plant
parts that appear enlarged, they will simply appear enlarged as a matter of course in drawing the plant.
For example, celery petioles will be thick simply because the petiole thickness parameter will be large. In
the case of storage organs the drawing algorithm will poorly reflect the model. Users will only be able to
find out the size of the edible items they harvest (which are invariably the storage organs) by looking at a
harvest report.
For the reproductive system, a similar system will be used to allocate biomass, with a parameter of the
fraction of photosynthate going to the reproductive system at maturity.
An example of some possible parameters is:
at HUI 0.2 0.5 0.95
Root 0.4 0.2 0.1
Flower/Fruit 0.0 0.1 0.5 (at water stress factor = 1.0)
Storage organ 0.0 0.05 0.3
Shoot 0.6 0.6 0.3
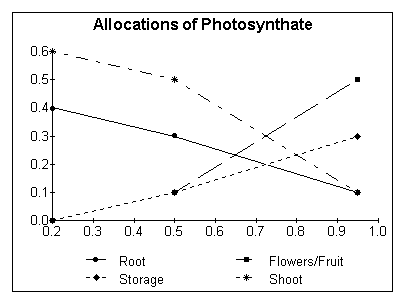
Possible allocation changes during the life of the plant
Separation of Simple Model From Complex Drawing Scheme
This is a very simple model that works as an extension to the simple root-shoot allocation model in
EPIC. In all respects the plant drawing algorithm reflects but does not affect the plant model,
which is very simple. The drawing scheme is a complex description of the underlying model state which
users can see, but does not load up the model with a lot of complexity we cannot test or verify.
Second Partitioning - Among Parts of the Shoot System
Biomass allocated to the shoots is further allocated to smaller parts of the shoot system -- meristems,
internodes, leaves, inflorescences, flowers, and fruit. The biomass allocation to these parts determines how
the plant appears on the screen. The purpose of this allocation is to give the user some idea of the health
and appearance of the plant based on its reactions to its environment and its genetics (parameters).
These are the main stages of flower and fruit development as they pertain to biomass partitioning:
period description switch to next period
floral initiation to flower timing of each flower's unfolding until inflorescence creates,
appearance and appearance according to number of
(each flower) flowers and days to inflorescence maturity
flower appearance to anthesis expansion and pollination of each number of days flowers take to reach
(each flower) flower full size plus number of days flower is open
anthesis to full size growth of each fruit number of days fruits take to get to full size
(each fruit)
ripening ripening of each fruit number of days fruits take to ripen
(each fruit)
Shoot Photosynthate Partitioning Among Vegetative Meristems
After new photosynthate is allocated to the root, shoot and fruit with no differentiation, shoot and
fruit photosynthate pools are divided again among the parts of each section. This second division is for
display only and has no effect on the model.
A plant's biomass object collection is made up of these object types:
 apical meristem - in
vegetative or reproductive mode apical meristem - in
vegetative or reproductive mode
axillary meristem - in
vegetative or reproductive mode
internode - a portion of
stem; attached to an internode above (or an apical meristem above) and an internode below, one or two
apical meristems, and one or two leaves (we are not handling whorled leaves)
leaf - attached to an
internode only
inflorescence -
attached to an internode on one side and to one or more flowers on the other
flower - attached to an
inflorescence on one side and one fruit on the other
fruit - attached to one
flower
When the plant first germinates, its biomass object collection consists of only one apical meristem with
zero associated biomass. It is assumed that the entire seed weight is used by the cotyledons and none is
allocated to the meristem. Photosynthesis at this point is presumably done by the cotyledons, but since
photosynthesis is not calculated with reference to leaves (and LAI has a minimum of 0.05), we will
assume the cotyledons are perfect and will not model them.
Each vegetative plant part (meristems in vegetative mode, internodes, leaves) makes a demand for
vegetative biomass based on either linear growth (meristems and internodes) or S-curved growth (leaves).
A traversing algorithm collects the total of these demands, computes the ratio of the total demand to the
total new shoot photosynthate, then gives each vegetative plant part a proportion of its demand based on
the ratio. There is no weighting by plant part or age, except in that older plant parts tend to demand more.
Allocation within the photosynthate allocated to shoots works by allocating photosynthate only to
meristems.
{bml PLDRAW1.WMF}
Vegetative biomass partitioning
Each active vegetative meristem provides a demand, which is the amount of biomass it needs (over
what it has already) to create an internode and one or two leaves. Each meristem gets only so many days
to create its internode and leaves. After the number of days is up, the meristem creates whatever size of
internode it can. The intial size of the internode and leaves is determined by how much photosynthate the
meristem was able to obtain in the number days it had to make them. After an internode of leaf is created,
it continues to demand biomass for some number of days until it can no longer grow.
After the shoot photosynthate is divided up, any meristems that reached their goal biomass create one
internode and one or two leaves. An axillary meristem that creates an internode turns into an apical
meristem and also creates one or more axillary meristems (one for each leaf created).
Internode enlargement
Though an internode makes biomass demands and grows in biomass after it is created, internode
enlargement (in length and width) depends partly on plant water uptake because much of internode
enlargement is due to water turgor. To simulate this in drawing, when an internode unit is created, it is
some percent of its full size. Usually at creation the percent of full size will be about 3% (since internodes
generally increase in size about 30 times the original size). Every day after creation, the internode unit
will attempt to move up some number of percentage points based on how many days it normally takes an
internode unit in a non-water-stressed plant to develop to full size. This number of days should be a
parameter for each cultivar because internode units can vary so much in size, from radish to corn. The
critical assumption here is that internode enlargement is linear from creation to full size. This assumption
is supported (roughly) by the literature. So say an internode unit starts at 3% of full size and normally
takes 5 days to reach full size. Each day it will attempt to enlarge by (100 - 3) / 5 = 19.4 percentage
points. The percentage which an internode unit actually increases is determined by this amount and the
water stress factor plant-wide. So if the water stress factor is 0.9, the internode unit will increase by 19.4 *
0.9 = 17.46 percentage points. In this way the history of the plant's water stress will be evident in the
sizes of the internodes. When an internode reaches a certain age, it can no longer expand and stays
forever at the size it has attained.
Note that this approach is purely for drawing, for reflecting the status of the plant model in the drawing.
There is no feedback to the model.
Leaf enlargement
Each leaf calculates its demand from an S curve of optimal growth, just as fruits do (see below). On
each day, the leaf demands enough photosynthate to take it to the point predicted for one day ahead on the
s curve. This will allocate the most photosynthate to middle-sized leaves and the least to new and old
leaves. The leaf draws its 3D object as a fraction of a full-sized leaf based on the ratio of its biomass to the
optimal biomass for a leaf.
Branching
If an axillary meristem does not branch, it is being affected by apical dominance. Apical dominance
is similar to "branching index" in the older plant drawing algorithm, in fact it is just the inverse of it.
How does an axillary meristem know if it should branch? There are two occasions on which an axillary
meristem can change.
1. On the day after the meristem is formed (the first full day it exists), the axillary meristem draws a
random number. If the number is below 1.0 - the current apical dominance, the meristem is active and
begins to demand photosynthate. If not, the meristem is non-active, and does nothing.
2. When the plant switches into reproductive mode, each non-active axillary meristem examines the
possibility of producing an inflorescence. If its stored random number falls below the floral index (the
probability that any non-active axillary meristem will produce an inflorescence), it begins to demand
enough photosynthate (from the reproductive pool) to produce an inflorescence. If the plant does not have
axillary flowers, this step does not happen.
Once the plant has reached the point of floral initiation, whether the plant has axillary or terminal flowers
comes into play.
1. If the plant has terminal flowers, each apical meristem considers its probability of continuing to
produce vegetative meristems versus creating reproductive meristems based on the floral index, which is
calculated from the number of apical inflorescences expected to be on the plant.
2. If the plant has axillary flowers, only axillary meristems will produce flowers, leaving apical meristems
free to continue producing internodes and leaves. Each axillary meristem that remains (has not branched)
will produce an inflorescence with the probability determined by the floral index. If it does not produce an
inflorescence it will simply do nothing for the rest of the plant's life. This decision is made only once,
when floral initiation occurs.
An axillary bud, from the period it is formed until the plant dies, has these three possibilities.
a. It will produce an internode soon after the bud is formed only if its branching probability is below the
branching index.
b. It will produce an inflorescence on the day the plant switches from vegetative to reproductive growth
only if its floral probability is below the floral index.
c. If it does not produce a vegetative unit or inflorescence, it will do nothing ever.
An apical bud has these two possiblities:
a. It will produce an internode soon after the bud is formed.
b. It will produce an inflorescence on the day the plant switches from vegetative to reproductive growth
only if its floral probability is below the floral index.
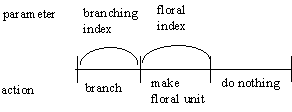
The decision to branch and how it is made
When you design the physical look of a plant, you manipulate the branching and floral indexes to
produce a plant with about the right amount of branches and about the right number of inflorescences.
Note: This model for apical dominance is not very good, and it will be improved in version 1.0.
Specifically we plan to simulate an amount of apical dominance hormone (or signal) which is passed as a
quantity from the apical meristems to the other meristems on the plant with a declining gradient away
from its source.
Reproductive biomass partitioning Among Reproductive Meristems and Fruit
Once floral initiation has occurred and some photosynthate is allocated to reproductive biomass, that
biomass must be allocated to flowers and fruits. The photosynthate is divided by demand among all
reproductive meristems and fruits.
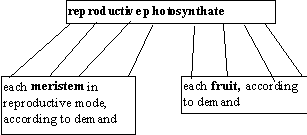
Reproductive biomass partitioning
Each meristem in a reproductive state demands how much photosynthate it needs to make one
inflorescence. Once the meristem has enough biomass to make one inflorescence, it creates the
inflorescence object, then is deleted. This does not work like the internode demand, in which only so
many days are available; a reproductive meristem will wait as long as it needs to.
Each fruit, like each leaf, calculates its demand from an S curve of optimal growth:
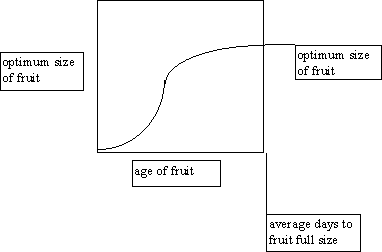
Fruit growth curve
On each day, the fruit demands enough photosynthate to take it to the point predicted for one day
ahead on the s curve. This will allocate the most photosynthate to middle-sized fruit and the least to new
and old fruits. The parameters needed here are the optimal biomass of a fruit and the number of days until
a fruit reaches full size.
Drawing Fruits
Fruits will be drawn with a scale that goes from some minimum percent of the full-sized fruit to the
full size of fruit (as determined in the plant parameter of fruit scale and the 3D object chosen for the fruit).
The percentage of maximum size to draw the fruit will be determined by the biomass of the fruit over the
optimum size of the fruit (which corresponds to the full-sized fruit 3D object).
Fruit Ripening
Fruits generally ripen some number of days after they were initiated, or at least attempt to, no matter
how large they are. So after the days to full size of the fruit, it will ripen, which means it will change color
(according to a parameter of fruit color after ripening). After a fruit enters the ripening stage it also will
stop demanding photosynthate.
Drawing Flowers
The inflorescence will control one or more flower objects. In the case of solitary flowers, the
inflorescence object doesn't really do anything; but in the case of real inflorescences, it encapsulates the
drawing behavior of the inflorescence. Each flower points to a fruit that it creates when it goes through
anthesis. Note that flower objects persist even after they are not drawn anymore. This is mostly for clarity.
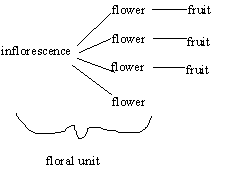
Architecture of inflorescences and flowers
The parameters for drawing the inflorescence mirror those used to draw the plant itself. Parameters
will be: number of flowers on the inflorescence (one for a single flower); branching index for the
inflorescence; peduncle length (the main stalk); internode length within the inflorescence; width of the
inflorescence stem; pedicel length (the stalk for each flower); and pedicel angle (between the internode
and the pedicel).
Different values of these parameters can create the most common inflorescence types: raceme, panicle and
umbel. We won't worry about cymes, which are sympodial racemes, because you can hardly see the things
anyway.
Flowers on inflorescences characteristically do not appear all at once, but one or two at a time. For this
reason the inflorescence creates one flower at a time, according to a parameter which tells how long the
inflorescence takes to create all the flowers. This parameter only makes sense if the inflorescence has
more than one flower.
The number of days between new flowers being created is floralUnitNumberOfFlowers /
floralUnitDaysToAllFlowersOpen. When the inflorescence creates a new flower, that flower does not
appear at full size; it takes several days to expand, like an internode, and demands biomass.
Flower expansion works like fruit expansion, only using linear growth. Flowers start as buds, with linear
growth as a bud, then switch to being open flowers, with continued linear growth, then open, then spend
some number of days reaching full size, then hesitate for a few days (parameter) before anthesis and the
creation of a fruit. Actually only one object is created for both the flower and fruit, since a flower becomes
one fruit.
As soon as the flower/fruit object reaches the fruit stage it begins drawing itself as a fruit and demanding
photosynthate as a fruit according to an S curve.
Separate Staminate and Pistillate Flowers
There are several vegetable plants that have separate male and female flowers (cucumber, squash,
corn), but in most of these the male flowers are small and inconspicuous and can be passed over in
drawing. Only in the corn plant is the male inflorescence large and important. For that reason all plants
must have two sets of information about inflorescences, one for male and one for female flowers. All the
information above must be duplicated for the two sets. For most plants the information for the male
flowers will not be needed.
Monoecious Plants
Only a few garden plants are monoecious; some spinach varieties and asparagus are two. In these
cases we can ignore the male plants and show only female plants, as if the male plants were around but
not to be seen. If more garden plants were of this type, it would be necessary to handle this, but for now I
think we should leave this alone.
Brussels Sprouts
A parameter for axillary bud scale is included so Brussels sprouts can have enlarged axillary buds.
Most plants will use a constant value for this parameter.
This software product, including its online help system and all printed documentation, is
Copyright (c) 1997 Paul D. Fernhout and Cynthia F. Kurtz All Rights Reserved.
Garden with Insight is a trademark of Paul D. Fernhout and Cynthia F. Kurtz.
| 